In Aging Cell, researchers have described the specific cell types that give exercise protective effects against motor nerve degeneration.
A different level of back problems
With aging, the prevalence of nerves connecting to muscle tissue dwindles, a phenomenon known as denervation [1]. This has been linked to a depopulation of the alpha motor neurons in the spinal cord [2]. Unless interventions are undertaken, the muscles themselves dwindle as a result [3], being gradually replaced by non-muscle fibrosis instead [4].
This fibrotic replacement is driven by meschenchymal fibroblasts [5] instead of the muscle fibroblasts that would replenish the normal tissue [6]. Fibroblasts have also been reported to secrete factors that encourage nerve regrowth [7], as have muscle stem cells [8].
While it is disputed, some work even suggests that, in middle age, human beings’ dwindling nerves are replaced by these natural processes [9] and that this results in more complex nerve centers than younger people have, particularly in people who exercise regularly [10]. While the accelerated decline caused by runaway aging processes leads to degradation without regeneration of the nervous tissue [11], intensive exercise has been found to efficiently protect against this [12].
The precise molecular mechanisms and cells involved, however, had not been fully described. This research focuses on muscle fibroblasts and stem cells in an effort to discover precisely what makes exercise such a powerful treatment for motor nerve degradation.
Cell type matters
In the first experiment, primary motor neurons were cultured from rat embryos, while muscle fibroblasts and stem cells were taken from human muscle biopsies. Despite the species difference, these cells were found to interact in a compatible way. Four of the human volunteers were young, four were old and sedentary, and six were old people who had exercised throughout their lives.
Muscle stem cells and fibroblasts were found to have starkly different gene expression profiles and stimulate the rat neurons in starkly different ways: a total of 11% of the neurons’ genes were expressed differently between the two groups, with roughly equal amounts of upregulation and downregulation. Considering the functions of these genes, culturing with fibroblasts seemed to encourage neural growth in a way that culturing with stem cells did not, including two genes that code for synapse transmission and a gene for the formation of new neurons (neurogenesis).
Culturing the rat neurons with conditioned media from these cell types, instead of directly, yielded somewhat similar results. Here, the researchers found that motor neurons are affected in complementary ways by each of these cell types, with fibroblasts still appearing to be more important for growth and development.
Lifelong exercisers have multiple advantages
The researchers then started comparing the cells of the different groups. Interestingly, they found that, while the young people were still better at exerting force than the older people and had slightly more lean mass, the older exercisers’ muscles appeared to be more efficient than younger people and older sedentary people. CAF, a key biomarker of neurological impairment, was not elevated in the older exercisers compred to the younger people, while older sedenary people had significantly elevated amounts.
Culturing the motor neurons with cells grown from older sedentary people had substantially negative effects: a full 53% more of the neurons survived after being cultured with cells derived from older exercisers versus their sedentary counterparts. The older exercisers’ cells even appeared to promote neuronal health more than those taken from younger people, although there was no statistically significant difference found with this small group.

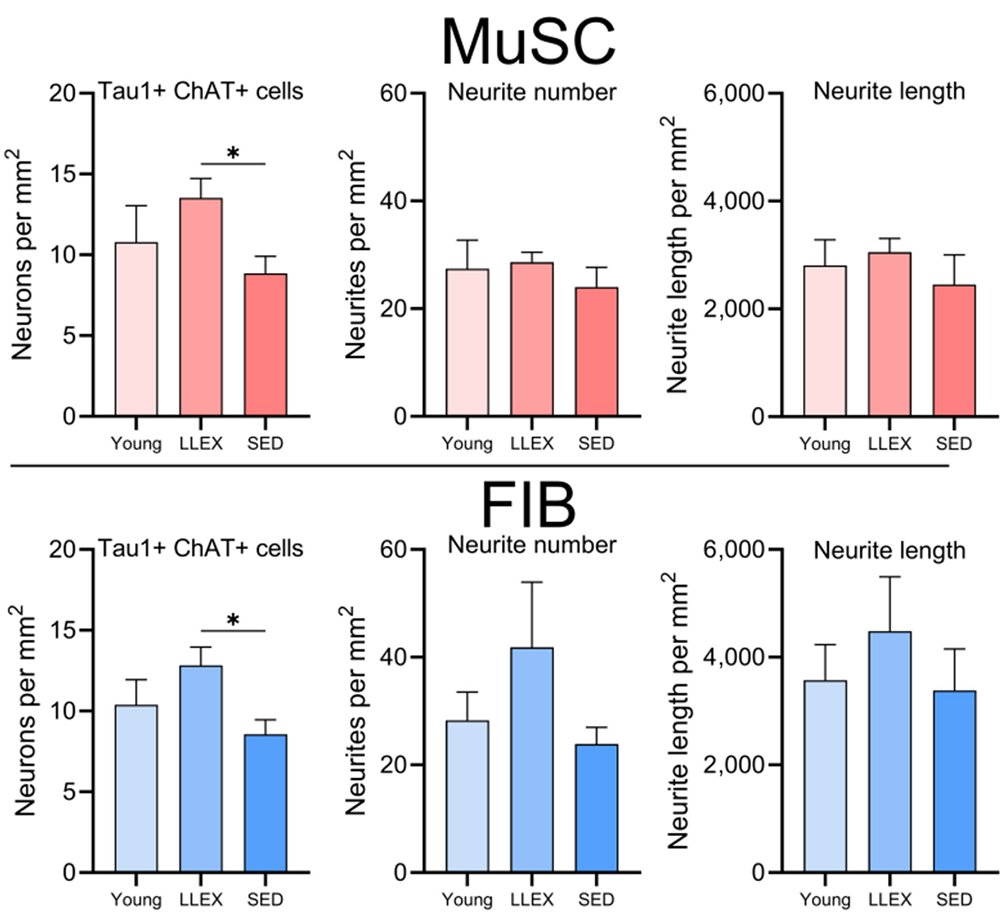
This was a relatively small study, and it did not delve into the precise molecular mechanisms involved in this communication between muscle fibroblasts, muscle stem cells, and motor neurons. However, it has clearly identified the cell types involved. An analysis of such factors as extracellular vesicles and other intercellular communication will be necessary to translate these findings into a useful treatment.
Despite its limitations, however, this study makes it absolutely clear that exercise is necessary for long-term muscle health and defense against motor neuron loss with age. It is unknown when life-changing biological interventions will enter the clinic; right now, according to this and many other studies, exercise remains one of the best treatments that is broadly available.
Literature
[1] Soendenbroe, C., Andersen, J. L., & Mackey, A. L. (2021). Muscle-nerve communication and the molecular assessment of human skeletal muscle denervation with aging. American Journal of Physiology-Cell Physiology, 321(2), C317-C329.
[2] McNeil, C. J., Doherty, T. J., Stashuk, D. W., & Rice, C. L. (2005). Motor unit number estimates in the tibialis anterior muscle of young, old, and very old men. Muscle & Nerve: Official Journal of the American Association of Electrodiagnostic Medicine, 31(4), 461-467.
[3] McPhee, J. S., Cameron, J., Maden-Wilkinson, T., Piasecki, M., Yap, M. H., Jones, D. A., & Degens, H. (2018). The contributions of fiber atrophy, fiber loss, in situ specific force, and voluntary activation to weakness in sarcopenia. The Journals of Gerontology: Series A, 73(10), 1287-1294.
[4] Madaro, L., Passafaro, M., Sala, D., Etxaniz, U., Lugarini, F., Proietti, D., … & Puri, P. L. (2018). Denervation-activated STAT3–IL-6 signalling in fibro-adipogenic progenitors promotes myofibres atrophy and fibrosis. Nature cell biology, 20(8), 917-927.
[5] Rebolledo, D. L., González, D., Faundez-Contreras, J., Contreras, O., Vio, C. P., Murphy-Ullrich, J. E., … & Brandan, E. (2019). Denervation-induced skeletal muscle fibrosis is mediated by CTGF/CCN2 independently of TGF-β. Matrix Biology, 82, 20-37.
[6] Wosczyna, M. N., & Rando, T. A. (2018). A muscle stem cell support group: coordinated cellular responses in muscle regeneration. Developmental cell, 46(2), 135-143.
[7] Theret, M., Rossi, F. M., & Contreras, O. (2021). Evolving roles of muscle-resident fibro-adipogenic progenitors in health, regeneration, neuromuscular disorders, and aging. Frontiers in Physiology, 12, 673404.
[8] Liu, W., Klose, A., Forman, S., Paris, N. D., Wei-LaPierre, L., Cortes-Lopez, M., … & Chakkalakal, J. V. (2017). Loss of adult skeletal muscle stem cells drives age-related neuromuscular junction degeneration. Elife, 6, e26464.
[9] Deschenes, M. R. (2011). Motor unit and neuromuscular junction remodeling with aging. Curr Aging Sci 4 (3): 209–220.
[10] Jones, E. J., Piasecki, J., Ireland, A., Stashuk, D. W., Atherton, P. J., Phillips, B. E., … & Piasecki, M. (2021). Lifelong exercise is associated with more homogeneous motor unit potential features across deep and superficial areas of vastus lateralis. GeroScience, 1-11.
[11] Snow, L. M., Mcloon, L. K., & Thompson, L. V. (2005). Adult and developmental myosin heavy chain isoforms in soleus muscle of aging Fischer Brown Norway rat. The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology: An Official Publication of the American Association of Anatomists, 286(1), 866-873.
[12] Soendenbroe, C., Heisterberg, M. F., Schjerling, P., Kjaer, M., Andersen, J. L., & Mackey, A. L. (2022). Human skeletal muscle acetylcholine receptor gene expression in elderly males performing heavy resistance exercise. American Journal of Physiology-Cell Physiology, 323(1), C159-C169.