A recent paper has reported on improving muscle function in aged mice through combined treatment of exercise and the inhibition of NNMT, an enzyme involved in NAD+ metabolism [1].
Age-related muscle loss
Muscle loss is a common problem in the aging population. Studies estimate that “after age 50, adults lose approximately 1% of their muscle mass and strength each year.” After turning 60, the annual muscle strength loss is even higher [2, 3].
Great effort has been invested in identifying possible treatments for age-related muscle decline and the resulting loss of skeletal muscle and strength (sarcopenia), with over 100 clinical trials in the past decade [4]. The authors of this study investigated nicotinamide N-methyltransferase (NNMT), which is involved in NAD+ metabolism and plays a role in the methylation of nicotinamide and DNA [5].
The group that conducted this study recently reported that in aged mice, the inhibition of NNMT increased contractile muscle function following injury [6]. This made them hypothesize that inhibiting NNMT in sedentary mice should improve muscle function similarly to exercise. On the molecular level, if the mechanisms behind exercise and NNMT inhibition are different, combining the treatments should have an additive effect on muscle function.
Increasing strength and running capacity
The researchers used aged (22-month-old) mice and divided them into sedentary and exercise groups. The exercise group was engaged in progressive weighted wheel running (PoWeR). Each group was further divided into two subgroups. One subgroup received a daily NNMT inhibitor for eight weeks, and the second was a control.
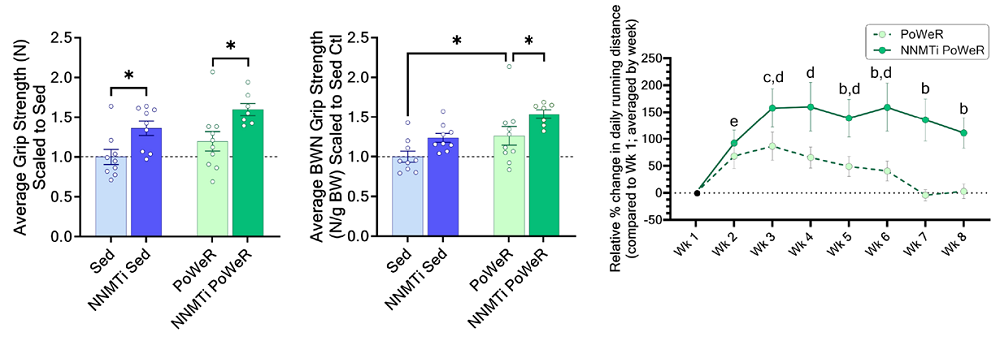
For their indicator of muscle function, the researchers chose forelimb grip strength. According to this measurement, both exercise and NNMT inhibition positively affected strength. Compared to untreated sedentary mice, the mice treated with an NNMT inhibitor had ~40% higher grip strength and the PoWeR group had a 20% increase in grip strength. Combining the NNMT inhibitor with exercise gave an even bigger effect than single treatments, and the researchers observed approximately 60% greater grip strength than the sedentary, untreated control group. A similar effect was observed when grip strength was normalized to body weight.
The results of this research agree with the results from a different group, which observed that NNMT inhibition ameliorated the loss of grip strength in aged mice.
The researchers also measured average weekly running distances in week one of the experiment and analyzed how the treatments influenced it. The NNMT inhibitor-treated PoWeR group’s running capacity increased, and this was maintained throughout the eight weeks of the experiment. In contrast, the inhibitor-untreated PoWeR group’s running capacity first increased; however, it was not increased as much as in the NNMT inhibitor-treated PoWeR group, and at week seven, it went back to the week one level.
According to the authors, this suggests “that NNMTi treatment improved tolerance to, or recovery from, intensive exercise.” The authors also note that mice treated with an NNMT inhibitor had reduced susceptibility to fatigue compared to controls.
Additionally, the researchers observed changes in muscle mass and composition. They observed that the mass of a few muscles, including the heart, increased. NNMT inhibitor treatment didn’t have such an effect on muscle mass.

On the molecular level
To understand the molecular mechanisms behind the observed effects of NNMT inhibitor treatment and exercise, the authors analyzed the proteins and metabolites produced in one of the large leg muscles of the mice.
The bulk analysis of the protein profiles of different treatment groups showed that the sedentary and exercise groups’ protein profiles were distinct. However, the protein profile of the NNMT inhibitor-treated sedentary group overlapped with that of the exercise group. Over three-quarters of proteins differentially regulated in the NNMT inhibitor-treated and exercise groups were regulated in the same direction (either increased or decreased). This suggests the involvement of many similar cellular processes.
The protein profile of the NNMT inhibitor-treated exercising group overlapped partly with that of the untreated exercise group. The researchers noted that only 15% of those proteins were regulated in the same direction (either increased or decreased) in both groups, suggesting that treating the exercising mice with an NNMT inhibitor might have modulated additional molecular processes and pathways.
Metabolite analysis identified a similar pattern to the protein analysis of the sedentary and exercise groups. Metabolites from those two cohorts created separate clusters, indicating great differences between these cohorts and exercise’s great impact on muscle metabolism.
Further analysis suggested that treating the sedentary mice with an NNMT inhibitor might have shifted the metabolic profile of these mice closer to that of the exercising mice.
The researchers also noted differences between the metabolic and protein profiles of the NNMT inhibitor-treated exercising cohorts. Contrary to its protein profile, the metabolic profile of that group was separate from that of other groups, suggesting that the combination of an NNMT inhibitor and exercise has a profound impact.
On a more granular level, when comparing the sedentary group treated with an NNMT inhibitor and the exercise group to sedentary controls, the researchers noted an increase in proteins involved in the protein production process (protein translation), which can explain the increased muscle weight. AMP, a low-energy signal that activates AMPK, was also increased in the mice in the exercise and NNMT inhibitor-treated groups. AMPK is a protein known for its energy-sensing properties, muscle hypertrophy, and protein translation [7, 8], linking those two observations.
Both interventions also reduced fat content in the muscle cells, which suggests improvements in muscle quality since high fat content in muscles “correlates with poorer muscle quality, functional performance, increased skeletal muscle insulin resistance.” [9, 10]
Possible treatment improvements
These researchers point out that using NNMT inhibitors could be an avenue for improving the current standard of care for age-associated loss of muscle mass and strength, which includes resistance or a combination of resistance and endurance exercise. However, bringing this to the clinic would require further testing in humans.
Literature
[1] Dimet-Wiley, A. L., Latham, C. M., Brightwell, C. R., Neelakantan, H., Keeble, A. R., Thomas, N. T., Noehren, H., Fry, C. S., & Watowich, S. J. (2024). Nicotinamide N-methyltransferase inhibition mimics and boosts exercise-mediated improvements in muscle function in aged mice. Scientific reports, 14(1), 15554.
[2] Beaudart, C., Locquet, M., Reginster, J. Y., Delandsheere, L., Petermans, J., & Bruyère, O. (2018). Quality of life in sarcopenia measured with the SarQoL®: impact of the use of different diagnosis definitions. Aging clinical and experimental research, 30(4), 307–313.
[3] von Haehling, S., Morley, J. E., & Anker, S. D. (2010). An overview of sarcopenia: facts and numbers on prevalence and clinical impact. Journal of cachexia, sarcopenia and muscle, 1(2), 129–133.
[4] Lo, J. H., U, K. P., Yiu, T., Ong, M. T., & Lee, W. Y. (2020). Sarcopenia: Current treatments and new regenerative therapeutic approaches. Journal of orthopaedic translation, 23, 38–52.
[5] Roberti, A., Fernández, A. F., & Fraga, M. F. (2021). Nicotinamide N-methyltransferase: At the crossroads between cellular metabolism and epigenetic regulation. Molecular metabolism, 45, 101165.
[6] Neelakantan, H., Brightwell, C. R., Graber, T. G., Maroto, R., Wang, H. L., McHardy, S. F., Papaconstantinou, J., Fry, C. S., & Watowich, S. J. (2019). Small molecule nicotinamide N-methyltransferase inhibitor activates senescent muscle stem cells and improves regenerative capacity of aged skeletal muscle. Biochemical pharmacology, 163, 481–492.
[7] Kong, L., Zhang, H., Lu, C., Shi, K., Huang, H., Zheng, Y., Wang, Y., Wang, D., Wang, H., & Huang, W. (2021). AICAR, an AMP-Activated Protein Kinase Activator, Ameliorates Acute Pancreatitis-Associated Liver Injury Partially Through Nrf2-Mediated Antioxidant Effects and Inhibition of NLRP3 Inflammasome Activation. Frontiers in pharmacology, 12, 724514.
[8] Attwaters, M., & Hughes, S. M. (2022). Cellular and molecular pathways controlling muscle size in response to exercise. The FEBS journal, 289(6), 1428–1456.
[9] Distefano, G., Standley, R. A., Zhang, X., Carnero, E. A., Yi, F., Cornnell, H. H., & Coen, P. M. (2018). Physical activity unveils the relationship between mitochondrial energetics, muscle quality, and physical function in older adults. Journal of cachexia, sarcopenia and muscle, 9(2), 279–294.
[10] Stefan, N., Kantartzis, K., Machann, J., Schick, F., Thamer, C., Rittig, K., Balletshofer, B., Machicao, F., Fritsche, A., & Häring, H. U. (2008). Identification and characterization of metabolically benign obesity in humans. Archives of internal medicine, 168(15), 1609–1616.