Researchers have outlined a key receptor and protein involved in the formation of cataracts, paving the way for potential treatments targeting them.
Oxidative stress and cataracts are closely related
Cataracts, which cloud the lenses of the eye, are one of the most common sources of age-related vision loss [1]. While they are treatable through surgery and artificial lens replacement, regenerative medicine is required to avoid surgical complications [2].
The aging of of lens epithelial cells (LECs), which normally surround the lens and keep it transparent [3], can cause cataracts as the cells suffer oxidative stress, protein misfolding, mishandling of calcium ions, and epigenetic alterations [4]. This also leads to these cells’ senescence and the iron-related cellular death known as ferroptosis [5].
Retinoic acid receptor-associated orphan receptors (RORs), most notably RORA, have been previously investigated in the context of eye diseases [6], including dry age-related macular degeneration [7] and oxygen-induced retinopathy [8]. RORA seems to play contradictory roles in different tissues: studies found that it is beneficial against oxidative stress in heart disease [9] and in Parkinson’s [10], but in wet AMD, it has been found to be harmful [11]. Therefore, these researchers decided to take a look at how RORA interacts with LECs.
Upregulation of this regulator worsens oxidative stress
These researchers created a model of cataracts by injecting rats with sodium selenite, a compound known to cause this condition. As expected, the injected rats suffered from severe lens damage a week later, including small voids and altered structure. This was accompanied by increased oxidative stress in LECs, increases in age-related biomarkers such as SA-β-gal, and markers of cellular death by apoptosis. RORA was similarly upregulated as well. Similar results were found in donated human tissues: RORs, most notably RORA, were upregulated in LECs that surrounded cataract lenses.
The researchers then turned to an in vitro experiment, culturing LECs in a medium rich in hydrogen peroxide in order to induce oxidative damage. As expected, markers of cellular senescence and apoptosis were increased in these cells along with oxidative stress, and RORA was increased as well.
However, this only reveals an association and does not prove that RORA has any kind of causative effect. To ascertain that, the researchers used one population of cells with RORA silenced and another population with RORA upregulated. RORA was found to be harmful rather than protective against oxidative stress: silencing it led to diminished stress markers, including markers related to apoptosis and cellular senescence. On the other hand, upregulating RORA in these cells increased these markers.
A downstream protein causes the damage
With the KEGG RNA database as their initial clue, the researchers found that the gene that encodes for prion protein (PRNP) was upregulated as a downstream consequence of RORA upregulation, a finding that was corroborated with PRNP upregulation in the donated cataract tissue, the in vitro cells exposed to hydrogen peroxide, and the rat model of cataracts. Another investigation using the JASPER database found that RORA has binding sites that directly led to increased expression of PRNP.
In vitro work found that PRNP was directly responsible for the increased oxidative stress associated with RORA in cells exposed to hydrogen peroxide. Upregulating PRNP neutralized the benefits of silencing RORA, increasing the associated senescence- and apoptosis-related biomarkers. This was confirmed by a rat experiment: silencing RORA in rats exposed to sodium selenite reduced their oxidative stress markers and cataract sizes, but upregulating PRNP reversed this effect.
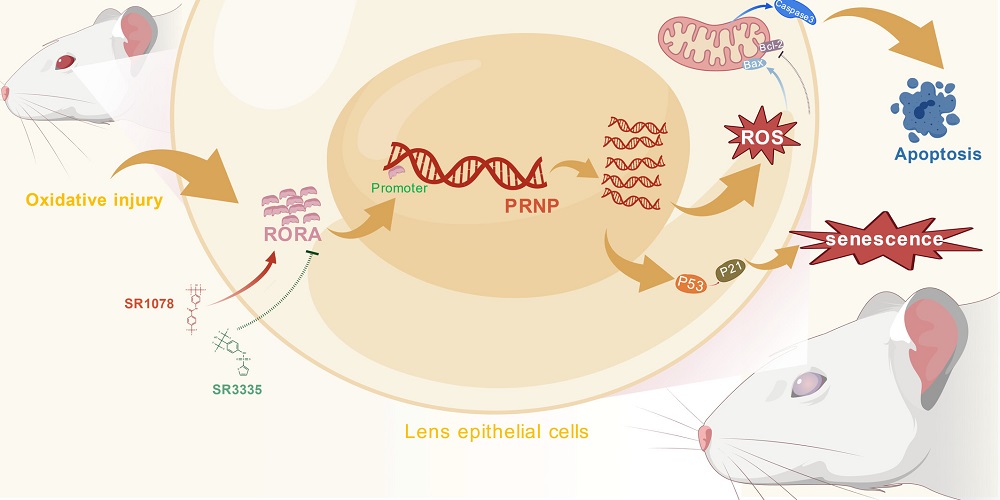
These researchers note the difficulties involved in targeting RORA itself, stating that the “efficient and targeted delivery of RORA inhibitors to lens tissue remains the core bottleneck limiting clinical translation of this strategy.” However, molecules that target PRNP or the RORA-PRNP axis, preventing RORA from activating PRNP, may be more feasible as clinical treatments. Future work will determine if such strategies can stop cataracts before they start and reduce the need for surgery.

Literature
[1] Liu, Y. C., Wilkins, M., Kim, T., Malyugin, B., & Mehta, J. S. (2017). Cataracts. The Lancet, 390(10094), 600-612.
[2] Lin, H., Ouyang, H., Zhu, J., Huang, S., Liu, Z., Chen, S., … & Liu, Y. (2016). Lens regeneration using endogenous stem cells with gain of visual function. Nature, 531(7594), 323-328.
[3] Mathias, R. T., White, T. W., & Gong, X. (2010). Lens gap junctions in growth, differentiation, and homeostasis. Physiological reviews, 90(1), 179-206.
[4] Periyasamy, P., & Shinohara, T. (2017). Age-related cataracts: Role of unfolded protein response, Ca2+ mobilization, epigenetic DNA modifications, and loss of Nrf2/Keap1 dependent cytoprotection. Progress in retinal and eye research, 60, 1-19.
[5] Wei, Z., Hao, C., Huangfu, J., Srinivasagan, R., Zhang, X., & Fan, X. (2021). Aging lens epithelium is susceptible to ferroptosis. Free Radical Biology and Medicine, 167, 94-108.
[6] Yemanyi, F., Bora, K., Blomfield, A. K., & Chen, J. (2023). Retinoic acid receptor-related orphan receptors (RORs) in eye development and disease. Retinal Degenerative Diseases XIX: Mechanisms and Experimental Therapy, 327-332.
[7] Akula, M., McNamee, S. M., Love, Z., Nasraty, N., Chan, N. P. M., Whalen, M., … & Haider, N. B. (2024). Retinoic acid related orphan receptor α is a genetic modifier that rescues retinal degeneration in a mouse model of Stargardt disease and Dry AMD. Gene Therapy, 31(7), 413-421.
[8] Sun, Y. E., Liu, C. H., Wang, Z., Meng, S. S., Burnim, S. B., SanGiovanni, J. P., … & Chen, J. (2017). RORα modulates semaphorin 3E transcription and neurovascular interaction in pathological retinal angiogenesis. The FASEB Journal, 31(10), 4492-4502.
[9] Xu, L., Su, Y., Zhao, Y., Sheng, X., Tong, R., Ying, X., … & Pu, J. (2019). Melatonin differentially regulates pathological and physiological cardiac hypertrophy: Crucial role of circadian nuclear receptor RORα signaling. Journal of Pineal Research, 67(2), e12579.
[10] Al-Zaid, F. S., Hurley, M. J., Dexter, D. T., & Gillies, G. E. (2023). Neuroprotective role for RORA in Parkinson’s disease revealed by analysis of post-mortem brain and a dopaminergic cell line. npj Parkinson’s Disease, 9(1), 119.
[11] Schaumberg, D. A., Chasman, D., Morrison, M. A., Adams, S. M., Guo, Q., Hunter, D. J., … & DeAngelis, M. M. (2010). Prospective study of common variants in the retinoic acid receptor–related orphan receptor α gene and risk of neovascular age-related macular degeneration. Archives of ophthalmology, 128(11), 1462-1471.